Перекисное окисление липидов — Википедия
Материал из Википедии — свободной энциклопедии
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 28 мая 2013; проверки требуют 9 правок. Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 28 мая 2013; проверки требуют 9 правок.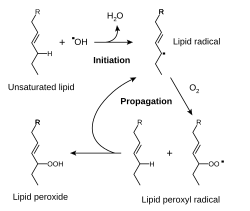
Перекисное окисление липидов (ПОЛ) — окислительная деградация липидов, происходящая, в основном, под действием свободных радикалов. Одно из главных последствий радиоактивного облучения.
В числе продуктов этого процесса — малондиальдегид и 4-гидроксиноненал.
Реакции биологического окисления сопровождаются образованием свободных радикалов — частиц, имеющих на внешней валентной орбитали неспаренный электрон. Это обусловливает высокую химическую активность этих радикалов. Например, они вступают в реакцию с ненасыщенными жирными кислотами мембран, нарушая их структуру. Антиоксиданты предотвращают свободнорадикальное окисление.
Через стадию перекисных производных ненасыщенных жирных кислот осуществляется биосинтез простагландинов и лейкотриенов, а тромбоксаны, оказывающие мощное влияние на адгезивно-агрегационные свойства форменных элементов крови и микроциркуляцию, сами являются гидроперекисями. Образование гидроперекисей холестерина — одно из звеньев в синтезе некоторых стероидных гормонов, в частности, прогестерона.[1][2]
Перекисное окисление липидов

Реакции переписного окисления липидов (ПОЛ) являются свободнорадикальными и по¬стоянно происходят в организме. Свободнора-дикальное окисление нарушает структуру мно¬гих молекул. В белках окисляются некоторые аминокислоты. В результате разрушается струк¬тура белков, между ними образуются ковалент-ные «сшивки», всё это активирует протеолити-ческие ферменты в клетке, гидролизующие повреждённые белки. Активные формы кисло¬рода легко нарушают и структуру ДНК. Неспе¬цифическое связывание Fe2+ молекулой ДНК облегчает образование гидроксильных радика¬лов, которые разрушают структуру азотистых оснований. Но наиболее подвержены действию активных форм кислорода жирные кислоты, содержащие двойные связи, расположенные через СН2-группу. Именно от этой СН2-группы свободный радикал (инициатор окисления) лег¬ко отнимает электрон, превращая липид, содер¬жащий эту кислоту, в свободный радикал.
ПОЛ — цепные реакции, обеспечивающие расширенное воспроизводство свободных ра¬дикалов, частиц, имеющих неспаренный элек¬трон, которые инициируют дальнейшее распро¬странение перекисного окисления.
В. ПОВРЕЖДЕНИЕ КЛЕТОК В РЕЗУЛЬТАТЕ
ПЕРЕКИСНОГО ОКИСЛЕНИЯ ЛИПИДОВ
Активные формы кислорода повреждают структуру ДНК, белков и различные мембран¬ные структуры клеток. В результате появления в гидрофобном слое мембран гидрофильных зон за счёт образования гидропероксидов жир¬ных кислот в клетки могут проникать вода, ионы натрия, кальция, что приводит к набуха¬нию клеток, органелл и их разрушению. Акти¬вация перекисного окисления характерна для многих заболеваний: дистрофии мышц (болезнь Дюшенна), болезни Паркинсона, при которых ПОЛ разрушает нервные клетки в стволовой части мозга, при атеросклерозе, развитии опу¬холей. Перекисное окисление активируется также в тканях, подвергшихся сначала ишемии, а затем реоксигенации, что происходит, напри¬мер, при спазме коронарных артерий и после¬дующем их расширении.
Такая же ситуация возникает при образова¬нии тромба в сосуде, питающем миокард. Формирование тромба приводит к окклюзии про¬света сосуда и развитию ишемии в соответству¬ющем участке миокарда (гипоксия ткани). Если принять быстрые лечебные меры по разрушению тромба, то в ткани восстанавливается снабже¬ние кислородом (реоксигенация). Показано, что в момент реоксигенации резко возрастает об¬разование активных форм кислорода, которые могут повреждать клетку. Таким образом, даже несмотря на быстрое восстановление кровооб¬ращения, в соответствующем участка миокарда происходит повреждение клеток за счёт актива¬ции перекисного окисления.
Изменение структуры тканей в результате
ПОЛ можно наблюдать на коже: с возрастом
увеличивается количество пигментных пятен
на коже, особенно на дорсальной поверхности
ладоней. Этот пигмент называют липофусцин,
представляющий собой смесь липидов и бел¬
ков, связанных между собой поперечными ко-
валентными связями и денатурированными в
результате взаимодействия с химически актив¬
ными группами продуктов ПОЛ. Этот пигмент
фагоцитируется, но не гидролизуется фермен¬
тами лизосом, и поэтому накапливается в клет¬
ках, нарушая их функции. °
ПОЛ происходит не только в живых организ¬мах, но и в продуктах питания, особенно при
Регуляция ПОЛ
Процессы ПОЛ усиливаются при избытке катехоламинов (стресс), гипоксии, ишемии, повышенном содержании активных форм О2, снижении антиоксидантной защиты, повышенном содержании ненасыщенных жирных кислот.
Биологическое значение ПОЛ
Модифицирует физико-химические свойства биомембран: изменяется проницаемость, активность мембранных ферментов.
Регулирует окислительное фосфорилирование.
Синтез ряда гормонов (стероидных), простагландинов.
Контроль клеточного деления.
Обмен ХС
Участвует в адаптации организма.
Повышение ПОЛ при патологии приводит к:
Разрушению, фрагментации клеточных мембран, повреждению и гибели клеток.
ПОЛ модифицирует ЛП, особенно ЛПНП. Они легче проникают в сосудистую стенку, лучше захватываются макрофагами, что ускоряет развитие атеросклероза.
Продукт ПОЛ малоновый диальдегид (МДА) — токсичен, канцерогенен, мутагенен.
ПОЛ ускоряет процесс старения организма.
Перекисное окисление липидов
Перекисному окислению липидов подвергаются полиненасыщенные ЖК, свободные или входящие в состав омыляемых липидов, при взаимодействии с активными формами кислорода.
Реакции переписного окисления липидов (ПОЛ) являются свободнорадикальными и по¬стоянно происходят в организме. Свободнора-дикальное окисление нарушает структуру мно¬гих молекул. В белках окисляются некоторые аминокислоты. В результате разрушается струк¬тура белков, между ними образуются ковалент-ные «сшивки», всё это активирует протеолити-ческие ферменты в клетке, гидролизующие повреждённые белки. Активные формы кисло¬рода легко нарушают и структуру ДНК. Неспе¬цифическое связывание Fe2+ молекулой ДНК облегчает образование гидроксильных радика¬лов, которые разрушают структуру азотистых оснований. Но наиболее подвержены действию активных форм кислорода жирные кислоты, содержащие двойные связи, расположенные через СН2-группу. Именно от этой СН2-группы свободный радикал (инициатор окисления) лег¬ко отнимает электрон, превращая липид, содер¬жащий эту кислоту, в свободный радикал.
ПОЛ — цепные реакции, обеспечивающие расширенное воспроизводство свободных ра¬дикалов, частиц, имеющих неспаренный элек¬трон, которые инициируют дальнейшее распро¬странение перекисного окисления.
В. ПОВРЕЖДЕНИЕ КЛЕТОК В РЕЗУЛЬТАТЕ
ПЕРЕКИСНОГО ОКИСЛЕНИЯ ЛИПИДОВ
Активные формы кислорода повреждают структуру ДНК, белков и различные мембран¬ные структуры клеток. В результате появления в гидрофобном слое мембран гидрофильных зон за счёт образования гидропероксидов жир¬ных кислот в клетки могут проникать вода, ионы натрия, кальция, что приводит к набуха¬нию клеток, органелл и их разрушению. Акти¬вация перекисного окисления характерна для многих заболеваний: дистрофии мышц (болезнь Дюшенна), болезни Паркинсона, при которых ПОЛ разрушает нервные клетки в стволовой части мозга, при атеросклерозе, развитии опу¬холей. Перекисное окисление активируется также в тканях, подвергшихся сначала ишемии, а затем реоксигенации, что происходит, напри¬мер, при спазме коронарных артерий и после¬дующем их расширении.
Такая же ситуация возникает при образова¬нии тромба в сосуде, питающем миокард. Формирование тромба приводит к окклюзии про¬света сосуда и развитию ишемии в соответству¬ющем участке миокарда (гипоксия ткани). Если принять быстрые лечебные меры по разрушению тромба, то в ткани восстанавливается снабже¬ние кислородом (реоксигенация). Показано, что в момент реоксигенации резко возрастает об¬разование активных форм кислорода, которые могут повреждать клетку. Таким образом, даже несмотря на быстрое восстановление кровооб¬ращения, в соответствующем участка миокарда происходит повреждение клеток за счёт актива¬ции перекисного окисления.
Изменение структуры тканей в результате
ПОЛ можно наблюдать на коже: с возрастом
увеличивается количество пигментных пятен
на коже, особенно на дорсальной поверхности
ладоней. Этот пигмент называют липофусцин,
представляющий собой смесь липидов и бел¬
ков, связанных между собой поперечными ко-
валентными связями и денатурированными в
результате взаимодействия с химически актив¬
ными группами продуктов ПОЛ. Этот пигмент
фагоцитируется, но не гидролизуется фермен¬
тами лизосом, и поэтому накапливается в клет¬
ках, нарушая их функции. °
ПОЛ происходит не только в живых организ¬мах, но и в продуктах питания, особенно при
Регуляция ПОЛ
Процессы ПОЛ усиливаются при избытке катехоламинов (стресс), гипоксии, ишемии, повышенном содержании активных форм О2, снижении антиоксидантной защиты, повышенном содержании ненасыщенных жирных кислот.
Биологическое значение ПОЛ
Модифицирует физико-химические свойства биомембран: изменяется проницаемость, активность мембранных ферментов.
Регулирует окислительное фосфорилирование.
Синтез ряда гормонов (стероидных), простагландинов.
Контроль клеточного деления.
Обмен ХС
Участвует в адаптации организма.
Повышение ПОЛ при патологии приводит к:
Разрушению, фрагментации клеточных мембран, повреждению и гибели клеток.
ПОЛ модифицирует ЛП, особенно ЛПНП. Они легче проникают в сосудистую стенку, лучше захватываются макрофагами, что ускоряет развитие атеросклероза.
Продукт ПОЛ малоновый диальдегид (МДА) — токсичен, канцерогенен, мутагенен.
ПОЛ ускоряет процесс старения организма.
— повреждение липидов — Биохимия
Среди первичных механизмов повреждения клеток при окислительном стрессе лидирует окисление жирнокислотных остатков в фосфолипидах мембран. Это снижает их гидрофобность и нарушает устойчивость мембран, изменяет работу мембраносвязанных ферментов, повышает проницаемость мембран для ионов.
Реакции взаимодействия свободных радикалов с жирными кислотами получили широкую известность в связи с их актуальностью в пищевой промышленности. Появление неприятного запаха и прогоркание продуктов – это является проявлением перекисного окисления липидов (ПОЛ).
Основным субстратом для свободно-радикальных реакций являются двойные связи полиненасыщенных жирных кислот. В клеточных мембранах полиненасыщенные жирные кислоты находятся в составе фосфолипидов и гликолипидов. Также большое количество фосфолипидов с полиненасыщенными жирными кислотами локализуется в оболочке липопротеинов высокой, низкой и очень низкой плотности, что имеет значение в патогенезе атеросклероза.
В результате свободнорадикального окисления жирных кислот образуются гидроперекиси и диеновые конъюгаты (первичные продукты), которые очень нестабильны. При участии металлов переменной валентности они быстро метаболизируют во вторичные (альдегиды и диальдегиды) и третичные (шиффовы основания) продукты перекисного окисления липидов.
Перекисное окисление липидов включает в себя несколько стадий:
- Инициация.
- Развитие.
- Разветвление.
- Обрыв цепи.
В момент инициации, например, гидроксил-радикалом атакуется метиленовая группа, расположенная между двойными связями, и выбивается атом водорода, восстанавливающий гидроксил-радикал до воды. Далее в жирной кислоте происходит перестановка двойной связи, смещение радикальной группы и взаимодействие ее с кислородом. В результате образуется липопероксильный радикал.
Дальнейшее взаимодействие полученного липопероксильного радикала с соседними жирными кислотами приводит к его нейтрализации и появлению новых липоперекисных радикалов, т.е. к развитию линейной цепной реакции с появлением новых окисленных жирных кислот.
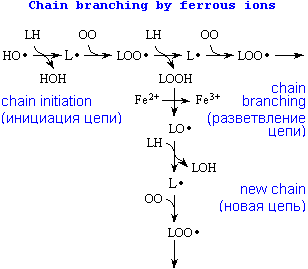
Развитие реакций перекисного окисления липидов
Кроме линейного развития, может происходить ветвление реакции за счет получения гидроперекисью электронов от каких-либо металлов или при воздействии излучения.
Разветвление и обрыв реакций перекисного окисления липидов
Обрыв цепной реакции происходит при взаимодействии радикалов друг с другом или в реакции с различными антиоксидантами, например, витамином Е, который отдаёт электроны, превращаясь при этом в довольно стабильную окисленную форму.
Продукты перекисного окисления липидов
Первичными продуктами ПОЛ являются гидроперекиси жирных кислот, они подвергаются дальнейшему распаду с образованием вторичных продуктов ПОЛ – различных спиртов, кетонов, альдегидов и диальдегидов, эпоксидов и других соединений.
Наиболее реакционноспособным из вторичных продуктов ПОЛ является малоновый диальдегид (МДА), который способен образовывать ковалентные связи с NH2-группами белков и иных молекул с образованием шиффовых оснований.
Схема реакций образования малонового диальдегида
Роль малонового диальдегида
Малоновый диальдегид (МДА), образующийся при перекисном окислении липидов, способен реагировать с ε-NH2-группами лизина или N-концевыми аминокислотами белков, с NH2-группами фосфолипидов и гликозаминов. МДА формирует мостики внутри молекул и между ними с образованием шиффовых оснований.
Роль малонового диальдегида в образовании сшивок между белками
Роль малонового диальдегида в образовании сшивок между белками и фосфолипидами
В конечном результате после окислительной атаки в белках появляются поперечные сшивки внутри одной молекулы, между разными белками, между белками и фосфолипидами. Из-за этого активность ферментативных белков изменяется, возможности структурных и сократительных белков падают, каналообразующие белки мембраны деформируются и проницаемость мембран возрастает, жизнеспособность и функционирование клетки уменьшаются.
Перекисное окисление липидов
Понятие ПОЛ объединяет все реакции неферментативного окисления полиненасыщенных ЖК, свободных или входящих в состав омыляемых липидов, протекающих по радикальному механизму. Реакции ПОЛ инициируются активными формами кислорода.


В результате появления в гидрофобном слое мембран гидрофильных зон за счёт образования гидропероксидов жирных кислот в клетки могут проникать вода, ионы натрия, кальция, что приводит к набуханию клеток, органелл и их разрушению.
Регуляция ПОЛ
Процессы ПОЛ усиливаются при избытке катехоламинов (стресс), гипоксии, ишемии (при реоксигенации), повышенном содержании активных форм О2, снижении антиоксидантной защиты, повышенном содержании ненасыщенных жирных кислот.
Активация ПОЛ происходит при воспалении и характерна для многих заболеваний: дистрофии мышц (болезнь Дюшенна), болезни Паркинсона, атеросклерозе, развитии опухолей.
Физиологическое значение реакций ПОЛ:
1. модифицируют физико-химические свойства биомембран: увеличивают их проницаемость.
2. регулируют активность мембранных ферментов, реакции окислительного фосфорилирования.
3. участвуют в контроле клеточного деления.
4. Участвует в адаптации организма.
Повышение активности ПОЛ приводит к:
1. разрушению, фрагментации клеточных мембран, повреждению и гибели клеток.
2. модификации ЛП, особенно ЛПНП. Они становятся «липкими», легче проникают в сосудистую стенку, хорошо захватываются макрофагами, что ускоряет развитие атеросклероза.
3. накоплению продуктов ПОЛ, многие из которых токсичны, канцерогенены и мутагенены (МДА).
4. ускорению процессов старения организма.
На коже с возрастом увеличивается количество пигментных пятен. Пигмент, вызывающий образование этих пятен, называется липофусцин. Он представляет собой смесь липидов и белков, связанных между собой поперечными ковалентными связями и денатурированными в результате взаимодействия с продуктами ПОЛ. Этот пигмент фагоцитируется, но не гидролизуется ферментами лизосом, и поэтому накапливается в клетках, нарушая их функции.
Дата добавления: 2016-07-27; просмотров: 1272;
Похожие статьи:
Что такое пол?
Наверное, никого не поставит в тупик вопрос, какого он пола, однако на вопрос «что нужно для того, чтобы родиться девочкой/мальчиком?» никто так и не даст исчерпывающего ответа. Американский психолог Джон Мани описал схему формирования пола у человека, состоящую из девяти этапов, или ступенек, которым и посвящена эта статья.
Хороший ты парень, Наташка, Наташка.
Зачем ты, Наташка,
Девчонкой на свет родилась?
(из детской песенки)
Долгое время считалось, что пол — это нечто монолитное и непреложное, определенное для каждого человека с самого момента оплодотворения яйцеклетки и неизменное в течение всей его жизни. Однако в XX веке американский психолог и сексолог Джон Мани предположил, что пол — это сложная многоуровневая система, формирующаяся многоступенчато в ходе онтогенеза под влиянием огромного числа всевозможных факторов.
Первая ступенька
Первой ступенькой становления пола человека является хромосомный пол — совокупность половых (X и Y) хромосом человека, возникающая в ходе оплодотворения яйцеклетки (рис. 1, слева). Так, у человека сочетание XX (гомогаметное) дает женский пол, а XY (гетерогаметное) — мужской. (Интересно, что не у всех животных это так: к примеру, у птиц, пресмыкающихся и бабочек гетерогаметным является женский пол.) Но, несмотря на столь простую схему определения хромосомного пола, и она дает сбои. Дело в том, что при образовании половых клеток может произойти нерасхождение половых хромосом в мейозе (таким образом, в одной клетке остаются две хромосомы; рис. 1, справа) или же «поломка» хромосомы. Такие грубые нарушения по большей части являются летальными (например, генотип Y0) или приводят к выкидышу на ранней стадии, однако некоторые из них жизнеспособны.

Рисунок 1. Хромосомы человека. Слева: Хромосомный набор мужчины. Справа: Схема нерасхождения Х-хромосом при оогенезе.
Синдром Шерешевского-Тернера — наличие одной X-хромосомы при неизменности состава соматических хромосом. Такая женщина имеет недоразвитые яичники (и поэтому бесплодна), а также комплекс врожденных аномалий — низкий рост, патологию шейных позвонков и т.д. (рис. 2). Частота синдрома Тернера — 1 на 700 тыс. новорожденных.

Рисунок 2. Фотография женщины с синдромом Шерешевского-Тернера и ее кариотип
Синдром Кляйнфельтера характеризуется наличием одной или нескольких дополнительных X-хромосом вдобавок к XY-кариотипу. Такие люди, как правило, имеют признаки обоих полов: высокий рост, маленький или среднего размера половой член, недоразвитые яички (сперматозоидов образуется мало, и они почти не жизнеспособные), но при этом округлые бедра и рост груди по женскому типу (рис. 3). Также у таких людей наблюдается снижение интеллекта, легочные расстройства и патология печени. Однако бывает, что статный молодой человек не имеет никаких признаков этого хромосомного нарушения, женится и лишь после неудачных попыток зачать ребенка узнает, что он не совсем мужчина. Частота рождения людей с синдромом Кляйнфельтера велика — 1 на 400 новорожденных.

Рисунок 3. Фотография больного с синдромом Кляйнфельтера и его кариотип
«Супермужской» и «суперженский» кариотипы — это появление дополнительной Y- или X-хромосомы, соответственно. «Супермужчины» имеют рост выше среднего и часто страдают бесплодием, умственной отсталостью и чересчур импульсивным поведением (считается даже, что именно такие мужчины составляют большую часть уголовников, но вызывающей доверие статистики на этот счет пока нет). «Суперженщины» же почти не имеют внешних аномалий; они способны к деторождению, однако у них раньше наступает менопауза, а также нередки умственные отклонения [1].
Вторая и третья ступеньки
Второй ступенькой на пути становления мужского и женского пола считается гонадный пол. Y-хромосома человека очень маленькая; она не может вместить в себя все гены, необходимые для развития гонад, поэтому они находятся и на Y-, и на X-, и даже на соматических хромосомах. «Дирижер мужской дифференцировки» — область SRY (sex-determining region Y), находящаяся на Y-хромосоме, — способна активировать гены других хромосом и контролировать развитие мужских гонад.
Развитие гонад [2] начинается на 4-й неделе внутриутробного развития человека, когда специальные зародышевые клетки — гонобласты — начинают мигрировать к будущим гонадам. Гены, регулируемые SRY-локусом, способствуют формированию яичек. Гонобласты образуют в них половые тяжи и начинают дифференцироваться в незрелые половые клетки (сперматогонии), покрытые эпителиальными клетками — предшественниками клеток Сертоли. Из мезенхимы, находящейся под зародышевым эпителием, дифференцируются клетки Лейдига, играющие важную роль в дальнейшем формировании внутренних и наружных половых органов.
Яичники женщин начинают формироваться на две недели позже, чем яички мужчин. Женские гонобласты, находящиеся в половых тяжах, начинают усиленно делиться, формируя овогонии — предшественники яйцеклеток, покрытые эпителиальными клетками, которые позже дифференцируются в фолликулярные клетки. Процесс формирования гамет заканчивается на 8 неделе внутриутробного развития, что знаменует собой прохождение организмом третьей ступеньки становления пола — гаметный пол.
Внутренние половые органы образуются из вольфовых (в случае мужского организма) и мюллеровых (в случае женского организма) протоков, которые изначально присутствуют у каждого зародыша независимо от пола. Вольфовы и мюллеровы протоки имеют различное происхождение и сначала являются частью мочевыводящей системы, однако в процессе образования почки они меняют квалификацию и принимают участие лишь в формировании половой системы.
Ступеньки 4–6
Четвертая ступенька — гормональный пол — тесно связана с предыдущей. Дело в том, что клетки Лейдига, находящиеся во вновь сформировавшемся яичке, начинают продуцировать тестостерон, а клетки-предшественники клеток Сертоли — мюллеров ингибирующий фактор (МИФ), которые вызывают рассасывание мюллерова протока и формирование внутренних и внешних половых органов по мужскому типу. Таким образом, у зародыша остается лишь вольфов проток, который в дальнейшем становится семяпроводом. Экспериментально доказано, что для инициации формирования внутренних и внешних органов женской половой системы не нужно никакого гормонального фактора; в отсутствие МИФа и тестостерона мюллеров проток сам становится маточными трубами, образуется матка, влагалище и внешние половые органы. К третьему месяцу беременности «ненужный» проток (вольфов в случае женского организма и мюллеров в случае мужского) дегенерирует, и эмбрион становится «однополым». Таким образом оказываются преодолены пятая и шестая ступеньки на пути к формированию пола — внешний и внутренний морфологический пол (рис. 4).

Рисунок 4. Формирование внешних и внутренних половых органов под действием МИФа и андрогенов (преимущественно тестостерона)
В ходе развития гаметного, гормонального и морфологического пола возможны нарушения, которые могут привести к истинному или ложному гермафродитизму.
Истинный гермафродитизм — чрезвычайно редкое явление (в литературе описано всего около 150 случаев), когда у человека есть и мужские, и женские половые органы. Как правило, хромосомный набор таких больных соответствует женскому кариотипу XX, однако в силу тех или иных причин вольфов канал не дегенерирует, а дает начало мужской половой системе. Также описаны случаи истинного гермафродитизма, возникшего из-за генетического мозаицизма или химеризма, когда некоторые клетки имеют мужской кариотип, а некоторые — женский.
Ложный гермафродитизм встречается достаточно часто (в среднем у 1 из 2000 новорожденных) и характеризуется аномалиями внешних половых органов при неполноценности внутренних и, следовательно, при дисрегулированном гормональном фоне.
Совсем недавно в журнале Placenta вышла статья о том, что материнский организм по-разному реагирует на плод в зависимости от пола последнего [3]. Так, ученые обнаружили несколько биомаркеров, уровень которых в крови матери выше, если плод женского пола. Этими биомаркерами оказались белки PLGF, s-Flt1 и PAI-2, играющие важную роль в развитии и функционировании плаценты и, следовательно, в адаптации кровообращения женского организма к беременности. Более того, если уровень этих биомаркеров в случае беременности мальчиком оказывается таким же, как у девочек, то велик риск различных осложнений беременности, таких как преэклампсия, преждевременные роды и рождение плода с недостаточной массой тела.
На этапе становления половых органов процесс формирования пола во внутреутробном развитии заканчивается. Однако есть еще несколько важных ступенек к тому, чтобы человек в полной мере был мужского (или женского) пола.
Седьмая ступенька
В первую очередь, не стоит забывать о седьмой ступеньке — пубертатном гормональном поле. Под контролем гипоталамуса и гипофиза гонады подростка начинают вырабатывать большое количество половых гормонов, как мужских (андрогенов), так и женских (эстрогенов), и именно от баланса их сигналов зависит формирование вторичных половых признаков молодого человека, а во многом и его самоидентификация.
Ступеньки 8 и 9
Восьмая и девятая ступеньки являются скорее психологическими составляющими пола, но они не менее важны, чем предыдущие семь. Речь идет о так называемом гражданском (паспортном) и социальном полах.
Паспортный пол — это то, кем предстает человек для людей, его окружающих: сначала перед акушеркой (поэтому одно из названий — акушерский пол), а позже — перед учителями, работодателями, то, как он записан в официальных документах — свидетельстве о рождении и паспорте.
С социальным полом немного сложнее. Эта ступенька определяется воспитанием человека, тем, какая роль ему приписывается в ближайшем окружении родственников и друзей. Одна из главных героинь популярного телесериала «Как я встретил вашу маму» — Робин Чарльз Щербатски — воспитывалась своим отцом как мальчик, и это наложило отпечаток на ее дальнейшую жизнь и поведение. Она любит виски, оружие и хоккей, не имеет подруг и прилагает большие усилия к тому, чтобы выглядеть женственной.
Основатель теории становления пола Джон Мани старался показать основополагающую роль социального окружения в этом процессе, но даже его ждало сокрушительное поражение: эксперимент провалился, а со временем вошел в топ-10 самых жестоких научных исследований. Тем не менее, никем не ставится под сомнение, что семья играет огромную роль в половой самоидентификации ребенка.
Однажды Джону Мани представился уникальный случай доказать всему миру, какую важную роль в определении пола ребенка играет то, кем — мальчиком или девочкой — его воспитывают. В одной американской семье родились мальчики-близнецы, один из которых вследствие неудачного обрезания потерял пенис. Обескураженные родители обратились к проф. Мани за советом, и тот рекомендовал им удалить мальчику половые железы и остатки пениса и в дальнейшем растить его как девочку. Так Брюс Реймер стал Брендой (рис. 5). Однако маленькая Бренда была странной девочкой: любила оружие и машинки, дралась с мальчишками в школе и даже отказывалась ходить в туалет сидя. В итоге Бренда наотрез отказалась делать последнюю пластическую операцию по моделированию внешних половых органов, и родителям пришлось рассказать ей о ее «мальчиковом» прошлом. После трех попыток суицида Бренда-Брюс отказался быть девочкой, прошел серию пластических операций по восстановлению мужских половых органов, женился и усыновил трех детей. Однако история Брюса-Бренды закончилась трагично: после смерти брата он покончил жизнь самоубийством, так и не сумев справиться с психологическими проблемами [4].

Рисунок 5. Брюс-Бренда Реймер — мальчик и девочка в одном лице
Так человек, начиная с первых часов своего существования, вынужден карабкаться со ступеньки на ступеньку, доказывая свою принадлежность к определенному полу. На каждом этапе возможны отклонения, и в конце концов то, кем будет ощущать себя ребенок, зависит от его воспитания, социальной роли, а отнюдь не только от хромосомного состава клеток его организма.
- O’Neil D. (2013). Sex chromosome abnormalities. Human chromosomal abnormalities: An Introduction to Common Gross Defects of Human Chromosomes;
- Silverman A.-J. Chapter 14: Gonadal development. In: Introduction to human development. 2004;
- Z.A. Brown, S. Schalekamp-Timmermans, H.W. Tiemeier, A. Hofman, V.W.V. Jaddoe, E.A.P. Steegers. (2014). Fetal sex specific differences in human placentation: A prospective cohort study. Placenta. 35, 359-364;
- Dr. Money and the Boy with No Penis. BBC;
- Асанов А.Ю., Демикова Н.С., Морозов С.А. Основы генетики и наследственные нарушения развития у детей. М.: Издательский центр «Академия», 2003. — 224 с.;
- Аскеров Н.А. Геномные болезни человека. Челябинск, 2016;
- Müllerian Agenesis. (2016). Stepwards;
- Agosto 22 – 1965. Nace David Reimer, un hombre forzado a ser mujer… quirurgicamente. (2015). Deep Politics;
- Сперматогенез: через тернии к звездам.
Как инфузория тетрахимена выбирает себе пол
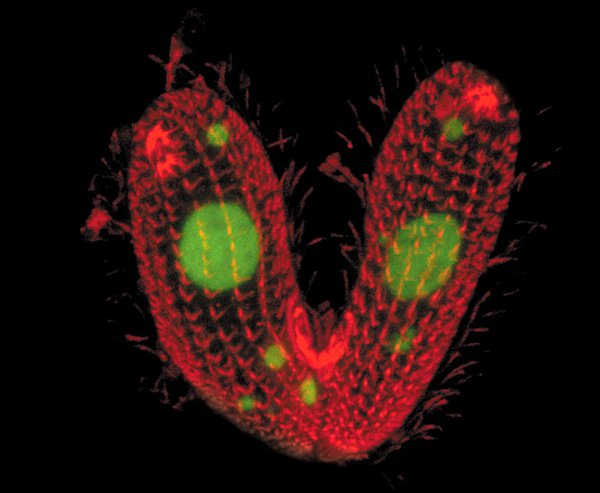
У инфузории Tetrahymena thermophila семь полов. Вступить в половой процесс она может с представителем любого пола, кроме своего. Клетки, получившиеся в результате полового процесса, не наследуют пол у своих «родителей» — они выбирают его случайным образом из нескольких вариантов. Коллектив ученых из США и Китая в деталях разобрался в этом аспекте личной жизни инфузорий. Известно, что у всех инфузорий два генома — один, рабочий, содержится в макроядре, а второй в неактивной форме хранится в микроядре и предназначен для обмена генетической информацией. Ученые выяснили, что в макроядре за пол отвечает пара расположенных по соседству генов. В микроядре тетрахимены есть все варианты таких пар, но в укороченной форме. После полового процесса одна из них достраивается путем «склеивания» частей разных генов, а все остальные уничтожаются. Удивительно, что выбор пары, по-видимому, происходит случайным образом, но при этом требует очень аккуратной сборки фрагментов ДНК.
В микромире «многополость», то есть наличие более двух полов, или типов спаривания, — не такое уж редкое явление. Оно весьма распространено среди грибов и простейших. Среди инфузорий тетрахимена совсем не чемпион по количеству полов — у представителя рода Euplotes их 12, а у инфузории Stylonychia mytilus, по некоторым данным, сто полов! В самом деле, при условии, что спариваться можно только с представителем другого пола, иметь всего два пола невыгодно — среди потенциальных половых партнеров остается только половина популяции. Если полов три, спариваться можно уже с 2/3 особей, и так далее. Таким образом, в погоне за генетическим разнообразием увеличивать количество полов может быть полезно.
Однако позволить себе это могут только одноклеточные микроорганизмы — ведь «пол» у них чаще всего определяется одним-двумя генами, кодирующими сигнальные белки (феромоны), позволяющие отличить «своих» от «чужих». У многоклеточных животных всё куда сложнее — для появления новых генетических комбинаций приходится создавать специальные половые клетки (гаметы), и чем сложнее устроена репродуктивная система, чем больше ресурсов тратится на воспроизводство, тем больше ограничений на количество вариантов. Да и зачем повышать генетическое разнообразие, если резервы многоклеточного организма позволяют противостоять неблагоприятным условиям. Возможно, поэтому более или менее высокоорганизованные животные обходятся двумя полами. Собственно, разобраться в этом вопросе — почему у одних полов много, а у других мало — и поможет изучение полового процесса у инфузорий.
Tetrahymena thermophila — один из модельных организмов, чей геном не так давно был расшифрован (см.: Число генов в геноме инфузории оказалось таким же, как у человека). О том, что у нее семь различных типов спаривания (то есть «полов»), обозначаемых как I–VII, знали еще с 50-х годов, однако никаких молекулярных подробностей о том, чем и как у нее определяется «пол», не было известно.
Напомним, что у инфузорий в клетке два ядра — одно из них (макронуклеус, или макроядро) содержит рабочий геном, который обеспечивает жизнедеятельность клетки, а второе (микронуклеус) является генеративным, то есть предназначено для обмена генетической информацией с другими клетками и содержит ДНК в неактивной форме. Обычно инфузории размножаются простым делением пополам (вегетативно), но если наступают голодные времена, они приступают к половому процессу. Для этого клетки частично объединяются (конъюгируют; см. рис. 1) и обмениваются половинками микронуклеусов. Новые микронуклеусы дают начало также новым макронуклеусам, в то время как старые разрушаются. Превращение генома микроядра в рабочий геном сопровождается обширными перестройками в ДНК. Более подробно этот процесс описан в новости Расшифрован геном инфузории Oxytricha trifallax — эукариотический геном из 16 000 хромосом, «Элементы», 23.03.2013.
Авторы статьи в журнале PLOS Biology своей работой положили начало изучению молекулярных механизмов личной жизни инфузорий. Они знали, что за пол у тетрахимены отвечает довольно большой участок (локус) в геноме макроядра, называемый mat-локус (от mating — спаривание). Для начала ученые проанализировали данные по экспрессии (то есть количество матричных РНК разных генов) в районе mat-локуса в поисках генов, которые неактивны в норме, когда инфузории размножаются вегетативно, но экспрессируются, когда инфузории голодают и хотят начать спариваться, и нашли пару кандидатов. Их назвали MTA и MTB. Выяснилось, что если эти гены удалить из генома, способность инфузорий к конъюгации сильно падает, то есть MTA и MTB действительно «гены спаривания». Подобная пара генов была обнаружена для каждого «пола». По всей видимости, эти гены кодируют белки. У каждого из предполагаемых белков нашли так называемый трансмембранный участок, то есть «якорь», который удерживает белок на мембране клетки. Трансмембранные участки оказались очень похожи у всех MT-белков, вне зависимости от пола инфузории.
Чтобы выяснить происхождение MT-генов в макроядре, стали искать их последовательности в геноме микроядра и обнаружили, что помимо MT-генов, характерных для актуального пола инфузории, геном микроядра содержит пары генов для всех вариантов пола, расположенные друг за другом в пределах одного локуса (рис. 2). Однако они оказались укороченными — отсутствовали последовательности, кодирующие те самые трансмембранные участки. Эти последовательности нашлись только у генов, расположенных по краям локуса. Возникают два вопроса: каким образом в макроядре собирается кассета, содержащая целые копии генов, характерные для данного пола, и куда деваются все остальные? (Под «кассетой» здесь понимается пара MT-генов, которые существуют как единое целое, то есть при перестройках генома так и остаются в паре.)
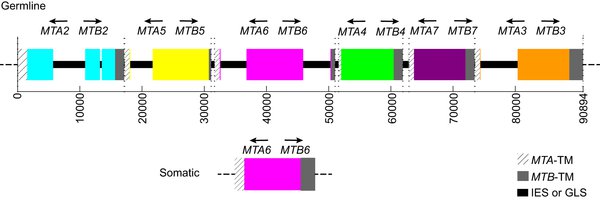
На второй вопрос оказалось ответить просто. Макрогеном инфузории проанализировали на наличие MT-генов, отвечающих за разные варианты пола, и нашли только гены, определяющие актуальный пол. Это означает, что все остальные наборы в процессе превращения микрогенома в макрогеном уничтожаются (в качестве альтернативы они могли бы просто быть «заглушены» при помощи химических модификаций ДНК и специальных белков, как это происходит у дрожжей (см. Mating of yeast). Таким образом, тетрахимена выбирает себе пол необратимо.
Дать ответ на первый вопрос оказалось куда сложнее. Определив последовательности, кодирующие трансмембранные участки у «родителей» и их потомства, ученые предположили, что сборка целостной кассеты происходит, когда недостающие участки ДНК с края локуса присоединяются к одной из укороченных кассет внутри локуса (рис. 3). Это может быть результатом либо гомологичной рекомбинации, то есть обмена похожими участками, либо просто акта разрезания-склеивания (так называемое негомологичное соединение концов). При этом требуется высокая точность сборки, так как присоединение происходит в кодирующей области гена и в случае ошибки синтезирующийся с такого гена белок будет нефункционален. В микроядре тетрахимены есть некоторое количество последовательностей, которые вырезаются при созревании макроядра (IES — internally eliminated sequences), однако они чаще всего расположены в некодирующих участках ДНК, и при их вырезании такая точность не соблюдается.
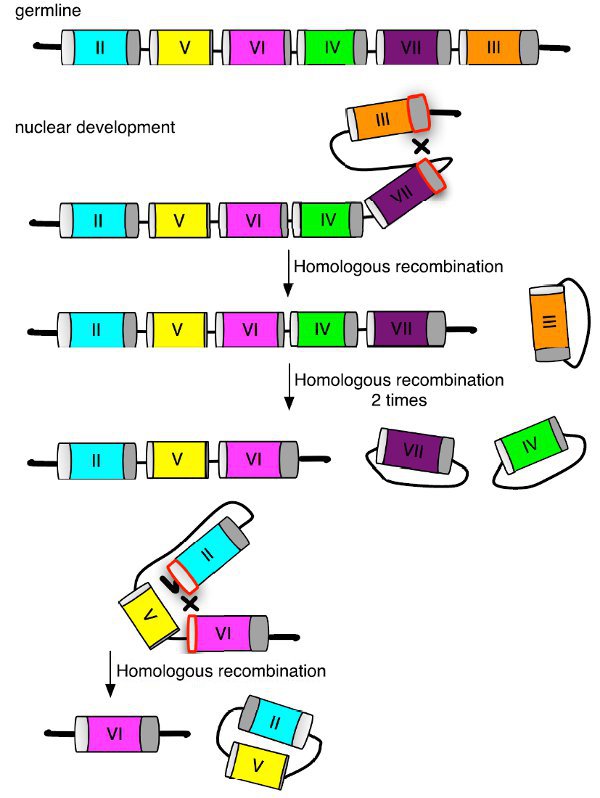
Непонятно, выпадает ли пара генов, которая будет определять будущий пол инфузории, при сборке случайно, как в рулетке, или перед процессом сборки она как-то выбирается и маркируется (например, путем химических модификаций ДНК), — это еще предстоит выяснить. Функции белков, кодируемых генами MTA и MTB, также пока неизвестны. Если это белки, лежащие на поверхности клетки, на что указывает наличие у них трансмембранного участка, наверняка они нужны для взаимного опознавания инфузорий. Самым интересным в этой истории остается вопрос, зачем тетрахимене целых семь полов. У ближайших ей видов количество полов варьируется от 3 до 9. Если каждый раз пол выбирается чисто случайно и никак не связан с полом «родителей», получается, что спаривание с представителем другого пола совсем не гарантирует притока свежих генов — партнер может оказаться ближайшим родственником. И почему тогда существует запрет на спаривание с представителем своего пола? Вероятно, в личной жизни инфузорий есть еще много нюансов, неизвестных исследователям.
Источник: Marcella D. Cervantes, Eileen P. Hamilton, Jie Xiong, Michael J. Lawson, Dongxia Yuan, Michalis Hadjithomas, Wei Miao, Eduardo Orias. Selecting One of Several Mating Types through Gene Segment Joining and Deletion in Tetrahymena thermophila // PLoS Biology. 2013. V. 11. P. e1001518. Doi:10.1371/journal.pbio.1001518. Статья в открытом доступе.
См. также:
Sujal S. Phadke, Rebecca A. Zufall. Rapid diversification of mating systems in ciliates // Biological Journal of the Linnean Society. 2009. V. 98. P. 187–197. Doi: 10.1111/j.1095-8312.2009.01250.x.
Дарья Спасская